Product Manager (Nutrition)
Stallen South Asia Pvt. Ltd.

Chief Operating Officer
Stallen South Asia Pvt. Ltd.

Product Manager (Therapeutics)
Stallen South Asia Pvt. Ltd.
1. Introduction
The poultry industry plays a crucial role in alleviating poverty by providing vast opportunities to millions of people across the country. According to the Government of India, the country ranks 3rd in the world for egg production and 5th for chicken meat production. The Indian poultry market was valued at INR 1,905.3 billion in 2022, with a growth rate of 6.77% for eggs (DADF) and 7%-8% for chicken meat (industry estimates). The annual per capita consumption is 101 eggs (DADF) and approximately 4.8-5 kg of chicken meat (based on the industry’s estimated placement of 500 crore day-old chicks).
However, there remains a significant gap in the consumption of poultry meat and eggs, primarily due to various challenges faced by the Indian poultry industry. One of the major challenges is the economic losses caused by infectious diseases. Among these, diseases caused by Mycoplasma species are particularly concerning (Marois et al., 2001). The primary pathogenic species are Mycoplasma gallisepticum (M. gallisepticum) and Mycoplasma synoviae (M. synoviae) (Umar et al., 2017). These Mycoplasma infections lead to substantial losses in the poultry industry by reducing egg production, stunting growth, and increasing condemnation rates at slaughterhouses. It has been reported that flocks affected by Mycoplasmosis may experience a 10-20% reduction in egg production (Bradbury and Morrow, 2008).
Stallen South Asia Pvt. Ltd. offers a wide range of products to control and prevent Mycoplasmosis which caused Chronic Respiratory Disease (CRD) and CCRD related conditions in poultry. The products include Forlutin 10% (Tiamulin Hydrogen Fumarate 10%), Doxatin (Tiamulin Hydrogen Fumarate 3.3%, Doxycycline HCl 2%) and Stylosin 100 (Tylosin Tartarate 10%) for prevention through feed additives and Forlutin 80% (Tiamulin Hydrogen Fumarate 80%) for treatment purpose through drinking water.
2. Mycoplasma in Poultry
Mycoplasmas are free-living, self-replicating bacteria characterized by having the smallest genome among bacteria, with a low guanine-cytosine (G+C) content ranging from 23% to 40% (Nicholas and Ayling, 2003). Unlike most bacteria, Mycoplasmas lack a cell wall, and their cell membranes are incorporated with sterols, which distinguishes them from other organisms. The absence of peptidoglycan makes them naturally resistant to antibiotics such as the beta-lactum antibiotics that target cell wall synthesis. Based on 16S rRNA analysis, Mycoplasma belongs to the phylum Firmicutes, class Mollicutes, and family Mycoplasmataceae (Ley, 2003; Ley, 2008). Among the 22 known species of Mycoplasma in birds, the four most common pathogenic species are Mycoplasma gallisepticum (M. gallisepticum), Mycoplasma synoviae (M. synoviae), Mycoplasma meleagridis (M. meleagridis), and Mycoplasma iowae (M. iowae). Of these, M. gallisepticum and M. synoviae are of particular importance due to their high prevalence in various types of poultry, with M. gallisepticum being considered the most pathogenic (Umar et al., 2017).
Sialoglycoprotein receptors in the respiratory epithelium are crucial for the attachment of Mycoplasma to epithelial cells, which is necessary for the initiation of the disease. This attachment process is mediated through cyto-adherence and is essential for Mycoplasma to evade the host’s innate defense mechanisms. Due to the absence of many metabolic pathways in Mycoplasma, these bacteria require close interaction with host cells for survival (Simecka et al.,1992).
3. Pathogenesis
3.1. Mycoplasma-Induced Cell Injury
Mycoplasma species can cause direct cell injury, although the exact mechanisms remain unclear. Mycoplasmosis leads to cell injury by depriving the host cells of essential nutrients, producing toxic substances, and altering host cell metabolites. Mycoplasma produces enzymes such as phospholipases, proteases, and nucleases, which damage host cell membranes and increase the risk of genetic alterations, potentially leading to autoimmune diseases (Bhandari and Asnani, 1989; Umar et al., 2017).
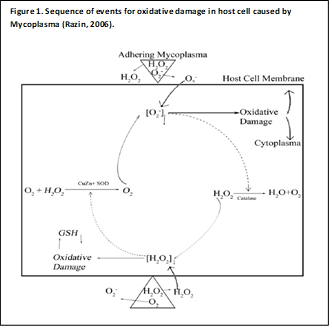
3.2. Role of Hydrogen Peroxide in Oxidative Damage
A critical factor in Mycoplasma-induced cell injury is the production of hydrogen peroxide, which significantly damages cell membranes and facilitates Mycoplasma adherence. The hydrogen peroxide released by Mycoplasma induces oxidative stress in host cells and can cause hemolysis. Catalase, an enzyme in host cells, converts hydrogen peroxide into nascent oxygen (O-2), further contributing to oxidative damage. To combat this, host cells produce antioxidant enzymes like glutathione (GSH) and superoxide dismutase (SOD), which help mitigate the oxidative stress caused by Mycoplasma (Razin et al., 1998).
4. Transmission
Mycoplasma is transmitted both vertically through eggs and horizontally through close contact, airborne droplets, and contaminated dust particles (Papazisi et al., 2002; Umar et al., 2017). The risk of transmission increases with the density of poultry populations in an area, often due to rapid expansion. This increased risk is one of the key reasons why maintaining Mycoplasma-free flocks is challenging (Lysnyansky et al., 2005)..
4.1. Mycoplasma gallisepticum
M. gallisepticum is the causative agent of Chronic Respiratory Disease (CRD) in chickens, with an incubation period of 16-21 days). The major clinical signs of CRD include gasping, respiratory rales, coughing, nasal discharge, and rhinitis. In some cases, M. gallisepticum can also lead to arthritis, salpingitis, conjunctivitis, and fatal encephalopathy. In egg-laying birds, the infection can cause a significant decrease in egg production and increased embryo mortality (Mukhtar et al., 2012).
4.2. Mycoplasma synoviae
M. synoviae is a significant pathogen in poultry worldwide, primarily due to the economic losses it causes, such as reduced egg production, growth retardation, and the condemnation of poultry meat at slaughterhouses. It typically causes infectious synovitis (a respiratory infection) in chickens, which can also result in subclinical infections. Currently, M. synoviae more commonly causes air sacculitis than infectious synovitis in chickens and turkeys. Air sacculitis can also occur because of co-infection with M. gallisepticum and E. coli. When the infection becomes systemic, it leads to inflammation of the synovial membranes of joints and tendon sheaths, resulting in synovitis, tenovaginitis, and bursitis (Kleven, 2008).
5. Tiamulin Hydrogen Fumarate
5.1. Chemical Composition
Tiamulin hydrogen fumarate (14-deoxy-14 [(2-diethylaminoethyl)-mercapto-acetoxy] mutilin hydrogen fumarate), known commercially as Forlutin10%, Forlutin 80% (Stallen South Asia. Pvt. Ltd), is a semisynthetic derivative of the diterpene antibiotic pleuromutilin. It is widely used in the treatment of airsacculitis, a condition primarily caused by Mycoplasma species. Animals infected with Mycoplasma spp. are more susceptible to various viral infections such as infectious bronchitis and Newcastle disease, as well as bacterial pathogens like Escherichia coli (causing coli septicemia). This susceptibility can lead to reduced growth rates, impaired feed conversion efficiency, and increased morbidity and mortality. Tiamulin is available as a crystalline powder, typically white to yellowish in color. It is commercially produced in several formulations: as a soluble powder with 45% tiamulin hydrogen fumarate in a lactose carrier, a 12.5% solution for inclusion in drinking water, and as a medicated feed premix available in strengths of 2%, 10%, and 80%. These formulations are widely available in most countries around the world.
5.2. Mechanism of Action of Tiamulin
Tiamulin exerts its antibacterial effects by binding to the rRNA within the peptidyl transferase slot on the ribosome. This binding interferes with the correct positioning of the CCA ends of tRNA, which is crucial for peptide transferase activity and subsequent protein production (Poulsen et al., 2001).
5.3. Microbial Resistance
Tiamulin has shown remarkable stability in terms of resistance development in Mycoplasma species. According to Valks and Burch (2002), Mycoplasma gallisepticum has exhibited almost no resistance development to tiamulin over the past 25 years. These findings are consistent with earlier research by Drews et al. (1975) which also indicated that tiamulin is generally a low inducer of resistance in Mycoplasma. Further studies by revealed that tiamulin targets the 50S subunit of the bacterial ribosome, particularly interacting at the peptidyl transferase center. They concluded that a mutation in the L3 protein of the ribosome, which points into the peptidyl transferase cleft, can lead to tiamulin resistance by altering the drug-binding site. Tiamulin has been shown to be compatible with tetracyclines in broilers. However, it is incompatible with nitrovin, a former growth promoter used in pigs and poultry. (Noa et al., 2000).
6. Tylosin Tartrate
6.1. Mechanism of action of Tylosin :
Tylosin tartrate (TT) is a macrolide antibiotic used in veterinary medicine and is extracted from the soil microbiome called Streptomyces fradiae . It is a bacteriostatic antibiotic that binds to the 50S subunit of the bacterial ribosome and inhibits bacterial protein synthesis. Stylosin-100 (Tylosin Phosphate 10%) is known to be strongly effective against Gram-positive and mycoplasma bacteria.
6.2. Use of Tylosin
Tylosin tartrate is effective in the treatment or prevention of chronic respiratory disease (CRD or CCRD) in poultry. It is particularly useful after vaccinations or during other stress conditions to prevent respiratory infections.
Conclusion
Stallen South Asia Pvt. Ltd. provides effective solutions for preventing and treatment of Mycoplasmosis in poultry caused, Chronic Respiratory Disease (CRD), and CCRD related conditions. Forlutin 10%, Doxatin & Stylosin-100 are feed additives that help prevent infections offers additional preventive support. For treatment, Forlutin 80% is administered through drinking water to manage active infections. This comprehensive product range supports overall poultry health and productivity.
References:
- “National Action Plan for Eggs and Poultry” prepared by DADF(2023-2024).
- BHANDARI, S., ASNANI, P.J. 1989. Characterization of phospholipase A2 of mycoplasma species. Folia Microbiologica, 34(4), 294-301.
- BRADBURY, J.M., MORROW, C. 2008. Avian mycoplasmas. Poultry Diseases, p. 220–234.
- Drews, J., A. Georgopoulos, G. Laber, E. Shütze, and J. Unger. 1975. Antimicrobial activities of 81.723 hfu, a new pleuromutilin derivative. Antimicrob. Agents Chemother. 7:507–516.
- KLEVEN, S.H. 2008. Control of avian mycoplasma infections in commercial poultry. Avian diseases, 52(3), 367-374. http://dx.doi.org/ 10.1637/8323-041808- Review.1
- LEY, D.H. 2003. Mycoplasma gallisepticum infection. In: Saif, Y.M., Barnes, H. J., Glisson J. R.,, Fadly, A. M., McDougald, L. R. and Swayne, D. E. (Eds.), Diseases of Poultry. Iowa State University Press, Ames, Iowa, USA: 722-744.
- Ley, D.L. 2008. Mycoplasmosis: Mycoplasma gallisepticuminfection. In Disease of Poultry, 11th Edition, pp. 807-834.
- LYSNYANSKY, I., GARCIA, M., LEVISOHN, S. 2005. Use of mgc2- polymerase chain reaction-restriction fragment length polymorphism for rapid differentiation between field isolates and vaccine strains of Mycoplasma gallisepticum in Israel. Avian diseases, 49(2), 238-45. http://dx.doi.org/10.1637/7285-10020R
- MAROIS, C., DUFOUR-GESBERT, F. & KEMPF, I.(2001). Molecular differentiation of Mycoplasma gallisepticum and Mycoplasma imitans strains by pulsed-field gel electrophoresis and random amplified polymorphic DNA. Journal of Veterinary Medicine, 48(9), 695-703.
- MUKHTAR, M., AWAIS, M. M., ANWAR, M. I., HUSSAIN, Z., BHATTI, N., ALI, S. 2012. Seroprevalence of Mycoplasma gallisepticum Among Commercial Layers in Faisalabad, Pakistan. Journal of Basic & Applied Sciences, 8, 183-186.
- NICHOLAS, R.A.J., AYLING, R.D. 2003. Mycoplasma bovis: disease, diagnosis, and control. Research in Veterinary Science, 74(2), 105-112
- Papazisi, L., Gorton, T. S., Kutish, G., Markham, P.F., Browning, G. F., Nguyen, D. K., Swartzell, S., Madan, A., Mahairas, G., Geary, S. J. 2003. The complete genome sequence of the avian pathogen Mycoplasma gallisepticum strain R(low). Microbiology, 149(9), 2307-2316. http://dx.doi.org/10.1099/mic.0.26427-0.
- Poulsen, S. M., M. Karlsson, L. B. Johansson, and B. Vester. 2001. The pleuromutilin drugs tiamulin and valnemulin bind to the RNA at the peptidyl transferase centre on the ribosome. Mol. Microbiol. 41:1091–1099.
- RAZIN, S., YOGEV, D., NAOT, Y. 1998. Molecular biology and pathogenicity of mycoplasmas. Microbiology and molecular biology reviews, 62(4), 1094- 1156.
- SIMECKA, J. W., DAVIS, J. K., DAVIDSON, M. K., ROSS, S.E., STÄDTLANDER, C.T. K.H., CASSELL, G.H. 1992. Mycoplasma diseases of animals. In Mycoplasmas: Molecular biology and pathogenesis, 391-415.
- UMAR, S., MUNIR, M.T., REHMAN, Z., SUBHAN, S., AZAM, T., SHAH, M.A.A. 2017. Mycoplasmosis in poultry: update on diagnosis and preventive measures. World’s Poultry Science Journal, 73(1), 1-12. http://dx.doi.org/10.1017/S0043933916000830.
- Valks, M., and D. G. S. Burch. 2002. Comparative activity and resistance development of tiamulin and other antimicrobials against avian mycoplasma. Page 200 in Proceedings of the XIIth World Veterinary Poultry Congress, Cairo, Egypt. Egyptian Veterinary Poultry Association, Cairo, Egypt.