
Chief Operating Officer
Stallen South Asia Pvt. Ltd.

Product Manager,
Stallen South Asia Pvt. Ltd.
Introduction
Respiratory disease remains a major limitation to productivity and welfare in commercial poultry production worldwide. Among bacterial pathogens, Mycoplasma gallisepticum (MG) and Mycoplasma synoviae (MS) are of particular importance due to their ability to establish chronic infections, interact synergistically with respiratory viruses, and persist within intensive production systems (1,2). The epidemiological and clinical impact of Mycoplasma infections has increased in recent decades as a consequence of production intensification, environmental stressors, viral evolution, and declining antimicrobial efficacy (3). These factors have shifted Mycoplasma infection from a discrete disease entity to a complex, multifactorial respiratory syndrome requiring integrated management and targeted therapeutic strategies.
Etiology and Biological Characteristics
MG and MS belong to the class Mollicutes, characterized by the absence of a cell wall, reduced genome size, limited metabolic capacity, and strict dependence on host-derived nutrients (4). The lack of a cell wall confers intrinsic resistance to β-lactam antimicrobials and allows close adherence to host cell membranes. High genetic plasticity and phase-variable surface lipoproteins facilitate antigenic variation and immune evasion, enabling long-term persistence within the host (4,5).
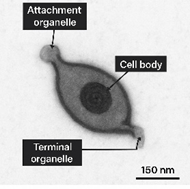
Pathogenesis and Lesion Development
Adherence to the respiratory epithelium is the critical initial step in Mycoplasma pathogenesis. MG and MS attach to ciliated epithelial cells of the trachea, lungs, air sacs, and conjunctiva via membrane-bound adhesins localized in specialized tip structures, including surface lipoproteins such as GapA and CrmA (4,6). Following colonization, Mycoplasmas induce ciliostasis, epithelial degeneration, and impairment of mucociliary clearance, promoting persistent infection and secondary bacterial invasion.
The host inflammatory response plays a central role in lesion development. Mycoplasma lipoproteins activate Toll-like receptor-2 pathways, stimulating epithelial cells and macrophages to release pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α (7). These responses result in mucosal edema, glandular hyperplasia, mucosal thickening, and accumulation of catarrhal to caseous exudate within the air sacs. Gross lesions commonly include airsacculitis, tracheitis, and, in complicated cases, fibrinous pericarditis and perihepatitis (2). MS may disseminate to synovial tissues, causing synovitis and lameness, and is strongly associated with eggshell apex abnormalities (EAA) in layers (8).
Transmission Dynamics and Epidemiology
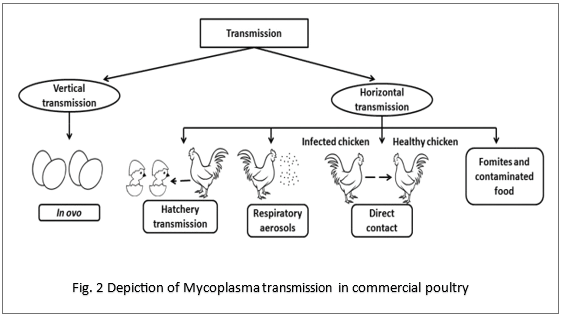
MG and MS persist in poultry populations through both vertical and horizontal transmission. Vertical transmission via infected eggs results in early colonization of chicks at hatch and contributes significantly to economic losses (9). Horizontal transmission occurs via aerosols, respiratory secretions, contaminated feed and water, and mechanical spread through personnel and equipment (2). Mycoplasmas may survive for several hours in dust and organic material, facilitating indirect transmission. Sub clinically infected carrier birds that intermittently shed organisms further complicate detection and control, particularly in multi-age production systems (1).
Disease Synergy and Environmental Influences
In commercial poultry, Mycoplasma infections rarely occur as single-agent diseases. MG infection predisposes the respiratory tract to enhanced replication of infectious bronchitis virus (IBV), resulting in increased air sac lesion severity, prolonged viral shedding, and higher viral loads (10). MS infection, particularly in association with nephropathogenic IBV strains, has been strongly linked to EAA, leading to downgraded eggs, reduced hatchability, and sustained production losses (8).
Synergistic interactions between Mycoplasma spp. and Newcastle disease virus (NDV) have also been reported, with even lentogenic NDV strains producing severe respiratory disease in the presence of Mycoplasma due to amplified epithelial damage and impaired mucosal immunity (11). Environmental stressors further exacerbate disease expression; ammonia impairs ciliary function, dust facilitates deep respiratory deposition of pathogens, and temperature stress suppresses mucosal immunity (3).
Antimicrobial Challenges in Mycoplasma Control
Antimicrobial therapy remains an important component of Mycoplasma control; however, increasing antimicrobial resistance has reduced the effectiveness of traditional agents. Rising minimum inhibitory concentrations (MICs) have been reported for macrolides, tetracyclines, and fluoroquinolones commonly used in poultry (12–14). The absence of a cell wall, high mutation rates, chronic persistence, and inappropriate antimicrobial use contribute to resistance development and therapeutic failure (12,13). In addition, Mycoplasmas adhere closely to host cell membranes and may occupy intracellular niches, limiting the efficacy of antimicrobials with poor tissue penetration.
Comparative Antimicrobial Activity Against Avian Mycoplasmas
Table 1 summarizes comparative in vitro activity (MIC₉₀ values) of antimicrobials commonly used in Mycoplasma control.
| Antimicrobial | MG MIC₉₀ (µg/mL) | MS MIC₉₀ (µg/mL) |
| Tylvalosin | 0.008–0.03 | 0.01–0.05 |
| Tylosin | 0.06–0.5 | 0.1–1.0 |
| Tilmicosin | 0.03–0.25 | 0.06–0.5 |
| Doxycycline | 0.25–2.0 | 0.5–4.0 |
| Enrofloxacin | 0.12–1.0 | 0.25–2.0 |
(12–15)
Pharmacological considerations for the Use of Tylvalosin
Tylvalosin is a semi-synthetic macrolide developed to address pharmacological limitations of earlier-generation antimicrobials. Structural modification of the tylosin molecule enhances lipophilicity, resulting in improved membrane permeability, intracellular accumulation, and tissue retention (16). Pharmacokinetic studies demonstrate rapid oral absorption and extensive tissue distribution, with lung concentrations four- to eight-fold higher than plasma levels (17).
In vitro studies consistently report low MIC₉₀ values against MG and MS (Table 1), indicating strong intrinsic activity against contemporary field isolates (15). Importantly, tylvalosin demonstrates superior intracellular penetration, including accumulation within macrophages, enabling effective targeting of intracellular Mycoplasma populations (16). Beyond antimicrobial activity, Tylvalosin exhibits immunomodulatory effects, including reduced production of TNF-α and IL-6 and decreased neutrophil infiltration, which may mitigate excessive inflammatory responses during Mycoplasma–viral synergistic infections (18).
Table 2. Pharmacokinetic and Pharmacodynamic Properties of Tylvalosin Compared with Other Antimicrobials
| Parameter | Tylvalosin | Tylosin | Tilmicosin | Doxycycline |
| Lung:plasma ratio | 4–8 : 1 | ~1–2 : 1 | ~3–4 : 1 | ~1 : 1 |
| Intracellular penetration | High | Low–moderate | Moderate | Moderate |
| Tissue retention | Prolonged | Short–moderate | Prolonged | Moderate |
| Anti-inflammatory effects | Demonstrated | Not reported | Not reported | Not reported |
(16–18)
Integration of Tylvalosin into Comprehensive Control Programs
While antimicrobial therapy alone cannot eradicate Mycoplasma, tylvalosin represents a rational option when integrated into comprehensive control programs including biosecurity, environmental optimization, vaccination against respiratory viruses, and routine monitoring using PCR and serology (1,2). Field studies in broilers, layers, and breeders demonstrate improved respiratory health, production performance, and reduced vertical transmission when tylvalosin is applied strategically (8,19).
Conclusion
Mycoplasma-associated respiratory disease remains a significant challenge in modern poultry production due to pathogen persistence, viral synergy, environmental stress, and antimicrobial resistance. Tylvalosin, with its favorable pharmacokinetic profile, strong activity against MG and MS, intracellular penetration, and immunomodulatory properties, addresses several limitations of traditional antimicrobials. When incorporated into integrated disease management strategies, tylvalosin contributes meaningfully to improved respiratory health and production outcomes in poultry.
References:
- Ley DH. Mycoplasma gallisepticum infection. Diseases of Poultry.
- Kleven SH. Control of avian mycoplasmosis. Avian Dis. 2008.
- Cook JKA et al. Infectious bronchitis and respiratory disease complexity. Avian Pathol.
- Razin S et al. Molecular biology of Mycoplasmas. Microbiol Mol Biol Rev. 1998.
- Gautier-Bouchardon AV. Antimicrobial resistance in avian Mycoplasma. Microbiol Spectr. 2018.
- Evans JD et al. Adhesion mechanisms of M. gallisepticum. Avian Dis.
- Muhlradt PF et al. Mycoplasma lipoproteins and immune activation. Infect Immun. 1997.
- Catania S et al. Mycoplasma synoviae and eggshell apex abnormalities. Avian Pathol. 2010.
- Carpenter TE et al. Economic impact of vertical MG transmission. Avian Dis. 1981.
- Evans RD. Synergism between MG and IBV. Avian Dis. 1998.
- Kleven SH. NDV–Mycoplasma interactions. Avian Dis. 2008.
- Hannan PC. Antibiotic resistance in Mycoplasma. J Vet Pharmacol Ther. 2000.
- Le Carrou J et al. MIC trends in avian Mycoplasma. Vet Microbiol. 2006.
- Lysnyansky I et al. In vitro susceptibility of avian Mycoplasmas. Avian Pathol. 2015.
- Gautier-Bouchardon AV et al. Resistance mechanisms in avian Mycoplasmas. Microbiol Spectr. 2018.
- Fischer AH et al. Macrolide intracellular pharmacodynamics. J Antimicrob Chemother. 2015.
- Taylor DJ. Macrolide pharmacokinetics in poultry. Vet J. 2012.
- Sirimalaisuwan A et al. Anti-inflammatory effects of tylvalosin. Res Vet Sci. 2013.
- McLaren J et al. Clinical efficacy of tylvalosin in broilers. Poultry Sci. 2017.